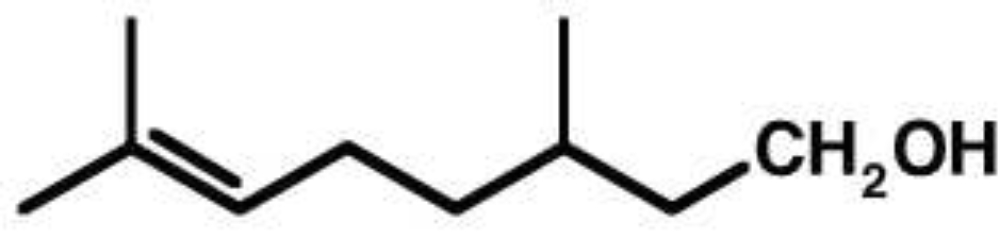

Terpene sind Verbindungen, deren Kohlenstoffgerüst auf Isopren (C5) als Grundkörper basiert und die daher seitenständige Methylgruppen besitzen. Monoterpene sind die einfachsten Terpene und bestehen aus zwei Isopreneinheiten. Man unterscheidet acyclische und cyclische Monoterpene. Alle Monoterpene sind mehr oder weniger angenehm riechende Aromaverbindungen (z. B. Citronellol [Zitrusaroma], Geraniol [Geranienaroma], Campher u. a.). Der typische Geruch vieler Pflanzen ist auf das Vorhandensein von (Mono-)Terpenen zurückzuführen. Weitere Isoprenverbindungen sind Carotinoide, Steroide, aber auch Kautschuk (Polyisopren). Citronellol (3,7-Dimethyl-6-octen-1-ol) und Citronellsäure (3,7-Dimethyl-2,6-octensäure) werden vielen Lebensmitteln als Aromastoffe zugesetzt. Monoterpene sind Bestandteil vieler Parfums. Interessanterweise riecht Citronellol nicht nur angenehm, sondern hat auch eine nützliche Wirkung als Insektenabwehrstoff und wird z.B. in Form von biologischen Repellents und Citronell-Kerzen vermarktet.
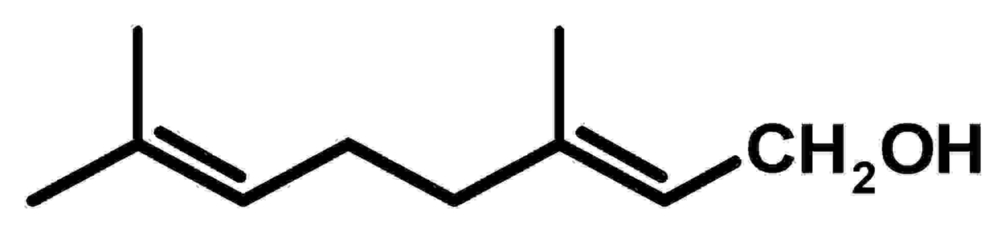

Vor einigen Jahren wurde entdeckt, dass Monoterpene wie Geraniol wichtige Effekte auf Säugerzellen haben und z.B. in menschlichen Zelllinien (auch Krebszelllinien) den programmierten Zelltod (Apoptose) auslösen können. Die dabei beteiligten Signalwege und biochemischen Transformationen der Terpene sind weitgehend unbekannt. Offenbar wurde die Wirkung von Monoterpenen auf den Säugerstoffwechsel bislang unterschätzt. Ein möglicher Grund dafür könnten unbekannte Stoffwechselwege sein, über die Monoterpene metabolisiert werden. In der Arbeitsgruppe Jendrossek wird daher der Stoffwechsel von Monoterpenen in Bakterien (Pseudomonas aeruginosa, Pseudomonas citronellolis) untersucht. Ein wichtiger Bestandteil des biochemischen „pathways“ ist der sogenannte Atu- und Liu-pathway (acyclic terpene utilization, Atu; leucine and isovalerate utilization, Liu).
Das Thema wurde 2014 abgeschlossen
Artikel:
Höschle, B., Jendrossek, D. 2005.
Utilization of geraniol is dependent on molybdenum in Pseudomonas aeruginosa: Evidence for different metabolic routes for oxidation of geraniol and citronellol.
Microbiology 151:2277-2283.
Höschle, B., Gnau, V., Jendrossek, D. 2005.
Methylcrotonyl-CoA carboxylase and geranyl-CoA carboxylase are involved in leucine/isovalerate utilisation (Liu) and in acyclic terpenes utilisation (Atu) and are encoded by liuA/liuC and atuC/atuF in Pseudomonas aeruginosa.
Microbiology: 151:3649-3656.
Förster-Fromme K., Höschle B., Mack C., Bott M., Armbruster W., Jendrossek D. 2006.
Identification of genes and proteins necessary for catabolism of acyclic terpenes and leucine/isovalerate in Pseudomonas aeruginosa.
Appl Environ Microbiol. 72:4819-28.
Förster-Fromme K, Jendrossek D. 2006.
Identification and characterization of the acyclic terpene utilization gene cluster of Pseudomonas citronellolis.
FEMS Microbiol Lett. 264(2):220-5.
Förster-Fromme, K., Chattopadhyay, A., and Jendrossek, D. 2008.
Biochemicalcharacterization of AtuD from Pseudomonas aeruginosa, the first member of a new subgroup of acyl-CoA dehydrogenases with specificity for citronellyl-CoA. Microbiology 154: 789-796.
Förster-Fromme, K., and Jendrossek, D. 2008.
Biochemical characterization of isovaleryl-CoA dehydrogenase (LiuA) of Pseudomonas aeruginosa and the importance of liu genes for a functional catabolic pathway of methyl-branched compounds.
FEMS Microbiol Lett. 286:78-84
Fromme, K.F.-F., Jendrossek, D. 2010.
AtuR is a repressorofacyclic terpene utilization (Atu) gene cluster expression and specifically binds to two 13 bp inverted repeat sequences of the atuA-atuR intergenic region. FEMS Microbiol. Lett. 308: 166-174
Chattopadhyay, A., Förster-Fromme, K., Jendrossek, D. 2010.
PQQ-dependent alcohol dehydrogenase (QEDH) of Pseudomonas aeruginosa is involved in catabolism of acyclic terpenes.
J. Bas. Microbiol. 50: 1-6
Fromme, K.F.-F., Jendrossek, D. 2010.
Catabolism of citronellol and related acyclic terpenoids in pseudomonads.
Minireview. Appl. Microbiol. Biotechnol. 87: 859–869
Siedenburg, G., and D. Jendrossek. 2011.
Squalene-hopene cyclases.
Appl Environ Microbiol 77:3905–3915.
Siedenburg, G., Jendrossek, D., Breuer, M., Juhl, B., Pleiss, J., Seitz, M., et al. (2012). Activation-independent cyclization of monoterpenoids. Applied and Environmental Microbiology, 78(4), 1055–1062. http://doi.org/10.1128/AEM.07059-11
Seitz, M., Klebensberger, J., Siebenhaller, S., Breuer, M., Siedenburg, G., Jendrossek, D., & Hauer, B. (2012). Substrate specificity of a novel squalene-hopene cyclase from Zymomonas mobilis. Journal of Molecular Catalysis B: Enzymatic, 84, 72–77. http://doi.org/10.1016/j.molcatb.2012.02.007
Siedenburg, G., Breuer, M., & Jendrossek, D. (2013). Prokaryotic squalene-hopene cyclases can be converted to citronellal cyclases by single amino acid exchange. Applied Microbiology and Biotechnology, 97(4), 1571–1580. http://doi.org/10.1007/s00253-012-4008-1
Poudel, N., Pfannstiel, J., Simon, O., Walter, N., Papageorgiou, A. C., & Jendrossek, D. (2015). The Pseudomonas aeruginosa Isohexenyl Glutaconyl Coenzyme A Hydratase (AtuE) Is Upregulated in Citronellate-Grown Cells and Belongs to the Crotonase Family. Applied and Environmental Microbiology, 81(19), 6558–6566. http://doi.org/10.1128/AEM.01686-15
Kontakt
